Gênese Estrutural: A Descoberta de uma Coordenação Única
A história da compreensão molecular do GHK-Cu começou quando pesquisadores demonstraram que a atividade biológica deste tripeptídeo não poderia ser explicada sem desvendar sua arquitetura estrutural — particularmente como três aminoácidos se organizam ao redor de um único íon cobre(II) para criar um complexo com propriedades biológicas extraordinariamente diversas. A química de coordenação do GHK-Cu determina sua estabilidade, sua capacidade de entrega de cobre, suas interações com receptores celulares e, fundamentalmente, sua habilidade de modular mais de 4.000 genes humanos.[1]
Esta análise estrutural detalhada destina-se a pesquisadores que buscam compreender a base química das propriedades biológicas do GHK-Cu. Para contextualização dos mecanismos biológicos possibilitados por esta estrutura, consulte nosso artigo sobre mecanismo de ação. Para implicações práticas das propriedades estruturais do GHK-Cu — particularmente sua sensibilidade à luz, dependência de pH e vias de degradação — veja nosso guia de manuseio e armazenamento.
Fundamentos Terapêuticos: Domínios de Aplicação Molecular
Regeneração Tecidual: Base Estrutural da Atividade
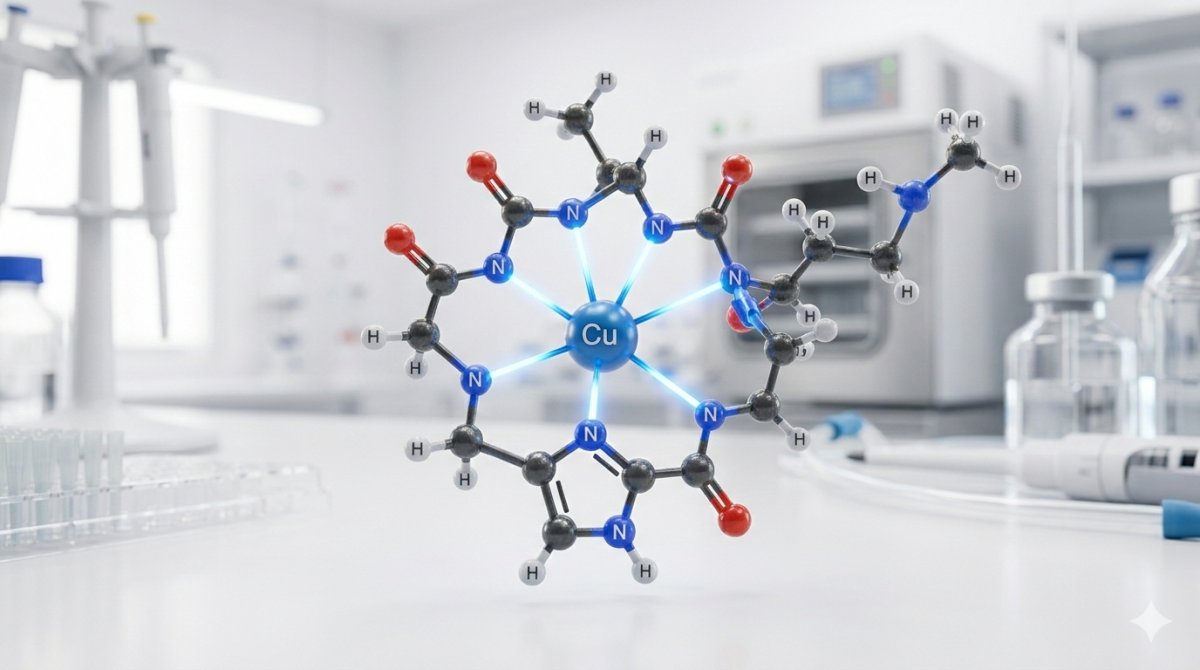
No domínio da regeneração tecidual, pesquisadores demonstraram que a estrutura do GHK-Cu é otimizada para modular a síntese de colágeno através de sua geometria de coordenação específica. O complexo consiste em três aminoácidos ligados por ligações peptídicas padrão: glicina (Gly, posição 1), L-histidina (His, posição 2) e L-lisina (Lys, posição 3). A fórmula molecular do peptídeo livre de cobre é C12H22N6O4, enquanto o complexo completo de cobre tem a fórmula C14H23CuN6O4 e peso molecular de 401,91 g/mol. O composto está registrado sob o número CAS 89030-95-5 e PubChem CID 73587.[2]
Cada resíduo contribui com propriedades distintas para o complexo regenerativo. A glicina, o menor aminoácido sem cadeia lateral, fornece flexibilidade conformacional no N-terminal e contribui tanto com seu grupo alfa-amino quanto com o nitrogênio da ligação peptídica para a coordenação do cobre. O anel imidazol da histidina é o local primário de ligação do cobre e a pedra angular estrutural de todo o complexo — átomos de nitrogênio do imidazol estão entre os doadores biológicos mais fortes para cobre(II), servindo como ligantes de cobre em numerosas metaloenzimas incluindo superóxido dismutase e citocromo c oxidase. A lisina fornece um grupo epsilon-amino positivamente carregado na terminação de sua cadeia lateral, contribuindo para a solubilidade aquosa do peptídeo, participando em interações eletrostáticas com superfícies biológicas aniônicas.[3]
Neuroproteção: Coordenação Molecular e Função Cerebral
No contexto neuroprotetivo, a sequência Gly-His-Lys não é arbitrária — ela é otimizada para ligação de cobre através de mecanismos específicos. A histidina na posição 2 é crítica porque posiciona o anel imidazol adjacente ao grupo amino N-terminal, criando uma cavidade de quelação idealmente dimensionada para cobre(II). Peptídeos com histidina na posição 3 ou mais distante do terminal exibem afinidade por cobre dramaticamente reduzida. A glicina na posição 1 é importante porque sua ausência de cadeia lateral minimiza a interferência estérica com a esfera de coordenação do cobre. A substituição da glicina por aminoácidos mais volumosos reduz a afinidade de ligação e altera a geometria do complexo resultante.[3]
Cicatrização: Geometria de Coordenação do Cobre(II)
Na esfera de coordenação primária relacionada à cicatrização, o íon cobre(II) no complexo GHK-Cu é coordenado por três átomos de nitrogênio doadores da cadeia peptídica. O primeiro doador é o nitrogênio Nδ1 ou Nε2 do anel imidazol da histidina (o doador individual mais forte no complexo). O segundo é o nitrogênio alfa-amino do resíduo de glicina no N-terminal. O terceiro é o nitrogênio da ligação peptídica desprotonado (nitrogênio da amida) entre glicina e histidina — uma coordenação que requer deslocamento do próton da amida e é característica de complexos cobre(II)-peptídeo fortes.[4]
Esta coordenação de três nitrogênios cria um quelato altamente estável com o íon cobre ligado dentro de dois anéis de cinco membros fusionados: um anel compreendendo Cu–NH₂(Gly)–CH₂–CO–N⁻ (o grupo amino da glicina através da ligação peptídica desprotonada) e um segundo anel envolvendo o nitrogênio imidazol da histidina. A geometria do anel de cinco membros é termodinamicamente favorável para coordenação de cobre(II), contribuindo para a constante de estabilidade notavelmente alta do complexo.[4]
Caracterização Estrutural: Métodos de Análise Avançada
Cristalografia: Revelando a Arquitetura Sólida
Análise cristalográfica de raios-X revelou que no estado sólido, a esfera de coordenação do cobre se estende além dos três doadores de nitrogênio peptídicos. O oxigênio carboxila do resíduo de lisina de uma molécula GHK-Cu vizinha ocupa uma posição de coordenação axial, criando uma geometria piramidal quadrada estendida (quatro doadores equatoriais mais um doador axial). Esta coordenação intermolecular produz uma estrutura de cadeia polimérica em cristais, onde unidades GHK-Cu adjacentes são ligadas através de pontes cobre-carboxilato.[4]
Em solução, a quarta posição equatorial é ocupada por uma molécula de água ou outro doador disponível, e a geometria flutua entre planar quadrada e piramidal quadrada dependendo da disponibilidade de ligantes axiais. Esta flexibilidade estrutural em solução pode ser funcionalmente significativa — permitindo que GHK-Cu adapte sua esfera de coordenação a diferentes ambientes celulares.[5]
Constante de Estabilidade: Quantificação da Afinidade
A estabilidade do complexo GHK-Cu é quantificada por sua constante de estabilidade log₁₀ (log K) de 16,44 — um valor excepcionalmente alto para um complexo tripeptídeo-cobre. Para comparação, o aminoácido histidina sozinho liga cobre(II) com um log K de aproximadamente 10,2, e a maioria dos complexos dipeptídeo-cobre exibem valores log K entre 8 e 14. A constante de estabilidade do GHK de 16,44 reflete a contribuição cooperativa de três doadores de nitrogênio em um arranjo geométrico ótimo, criando uma cavidade de quelação termodinamicamente pré-organizada para ligação de cobre(II).[3]
Esta alta afinidade tem implicações biológicas importantes. Em concentrações fisiológicas de cobre, o equilíbrio favorece fortemente a formação do complexo — essencialmente todo GHK na presença de cobre(II) suficiente existirá como GHK-Cu. No entanto, a afinidade não é tão extrema a ponto de impedir a liberação de cobre para aceptores biológicos de maior afinidade como albumina, ceruloplasmina ou chaperonas de cobre intracelulares.[1]
Complexos Binários e Ternários: Variações Estruturais
Complexo Binário: A Forma Fundamental GHK-Cu
A espécie GHK-cobre mais simples é o complexo binário 1:1 (GHK-Cu), no qual uma molécula de peptídeo coordena um íon cobre(II) através dos três doadores de nitrogênio descritos. Esta espécie predomina em pH fisiológico (7,4) quando GHK e cobre estão presentes em proporções equimolares ou próximas. O complexo binário é a forma mais extensivamente caracterizada espectroscopicamente e é a espécie mais comumente implicada pela designação "GHK-Cu" na literatura de pesquisa.[5]
Complexo Ternário: Coordenação Dupla (GHK)₂-Cu
Quando GHK está presente em excesso relativo ao cobre, um complexo ternário 2:1 pode se formar — designado (GHK)₂-Cu — no qual duas moléculas de peptídeo GHK coordenam o mesmo íon cobre(II). Nesta espécie, cada molécula de peptídeo contribui com seu nitrogênio imidazol da histidina e um ou dois doadores adicionais, com o cobre coordenado em uma geometria distorcida que acomoda ligantes de ambas as cadeias peptídicas.[5]
A relevância biológica do complexo ternário é uma área ativa de investigação. Estudos sobre penetração cutânea demonstraram que (GHK)₂-Cu pode atravessar o estrato córneo, sugerindo que tanto as formas binárias quanto ternárias podem contribuir para os efeitos tópicos do GHK-Cu.[6]
Comportamento Dependente de pH: Estabilidade Molecular
Coordenação em Função do pH
A química de coordenação do GHK-Cu é fortemente dependente do pH, uma propriedade com implicações práticas diretas para manuseio, formulação e desenho experimental. Em valores de pH ácidos (abaixo de aproximadamente 4,5), a protonação dos doadores de nitrogênio amino e imidazol interrompe a coordenação do cobre, levando à dissociação do complexo e liberação de íons cobre(II) livres. O cobre livre não está mais quelado em uma forma biologicamente controlada e pode catalisar geração de radicais tipo Fenton, contribuindo para degradação oxidativa do peptídeo.[3]
Em pH levemente ácido a neutro (5,0–7,4), os três doadores de nitrogênio são progressivamente desprotonados e a coordenação do cobre é estável. Esta faixa representa a janela funcional de pH para GHK-Cu e corresponde à faixa de estabilidade ótima para manuseio e armazenamento. O complexo atinge estabilidade máxima em torno de pH 7,4 (pH fisiológico), onde todos os três doadores de nitrogênio estão completamente engajados com o centro de cobre.[3]
Em pH alcalino (acima de 8,5), reações competitivas podem ocorrer, incluindo formação de hidróxido de cobre e potencial hidrólise da cadeia peptídica. Embora o complexo permaneça largamente intacto em pH levemente alcalino, condições fortemente básicas devem ser evitadas em formulações de pesquisa.
Caracterização Espectroscópica: Técnicas de Identificação
Espectroscopia UV-Visível: A Assinatura Azul
A característica mais imediatamente reconhecível do GHK-Cu adequadamente formado é sua cor azul real distintiva, decorrente de transições eletrônicas d-d do íon cobre(II) dentro de sua esfera de coordenação de doadores de nitrogênio. O espectro de absorção UV-visível exibe uma banda de absorção ampla centrada próximo a 600 nm (a banda de transição d-d característica de cobre(II) em um campo de ligante rico em nitrogênio) e uma banda de transferência de carga mais intensa na região UV próxima. Esta cor azul serve como um indicador prático de controle de qualidade: GHK-Cu adequadamente reconstituído deve produzir uma solução azul clara, e desvios — verde (indicando dissociação parcial do cobre), marrom (indicando oxidação) ou incolor (indicando perda completa do cobre) — sinalizam degradação.[4]
Ressonância Paramagnética Eletrônica (EPR)
A espectroscopia EPR fornece informações detalhadas sobre o ambiente de coordenação do cobre em GHK-Cu. Cobre(II) é um íon paramagnético d⁹, e seu espectro EPR é sensível ao número, tipo e geometria dos ligantes coordenantes. GHK-Cu exibe parâmetros EPR (valores g e constantes de acoplamento hiperfino) consistentes com uma geometria de coordenação planar quadrada ou tetragonalmente distorcida com doadores predominantemente de nitrogênio — confirmando o modelo de coordenação de três nitrogênios derivado de dados cristalográficos.[5]
Ressonância Magnética Nuclear (RMN)
Embora cobre(II) seja paramagnético e amplie sinais de RMN próximos (tornando difícil a observação direta de núcleos ligados ao cobre), a espectroscopia RMN de análogos diamagnéticos de zinco(II) e comparação de espectros de GHK livre de cobre com GHK-Cu forneceram informações sobre mudanças conformacionais induzidas pela ligação do metal. Estes estudos confirmam que a coordenação do cobre restringe a cadeia peptídica, reduzindo a flexibilidade conformacional do segmento Gly-His.[5]
Relações Estrutura-Atividade: Fundamentos Moleculares
O Papel Essencial do Cobre
Estudos comparativos entre GHK-Cu complexado com cobre e o peptídeo GHK livre de cobre demonstram consistentemente que o complexo completo exibe atividade biológica substancialmente maior em praticamente todos os endpoints medidos. Esta dependência do cobre é mecanisticamente lógica: a atividade antioxidante tipo SOD requer a capacidade de ciclagem redox do centro de cobre (Cu²⁺ ↔ Cu⁺); a entrega de cobre para metaloenzimas requer cobre biodisponível; e as restrições conformacionais impostas pelo cobre na cadeia peptídica podem ser necessárias para interações produtivas com receptores.[1]
Modificações de Sequência e Estudos com Análogos
Estudos de relação estrutura-atividade usando análogos de GHK iluminaram quais características estruturais são essenciais para a função biológica. A substituição da histidina por aminoácidos não coordenantes abole a maior parte da atividade biológica, confirmando a centralidade da coordenação imidazol-cobre. A substituição da glicina na posição 1 por aminoácidos maiores reduz a afinidade pelo cobre e atenua a atividade, demonstrando a importância da interferência estérica mínima no local de coordenação N-terminal.[3]
O Motivo ATCUN: Classificação Estrutural
GHK pertence a uma classe estrutural mais ampla conhecida como peptídeos do motivo ATCUN (ligação amino-terminal de cobre e níquel) — sequências com a fórmula geral X-X-His (onde X é qualquer aminoácido), que coordenam cobre(II) através do nitrogênio amino N-terminal, do nitrogênio da ligação peptídica desprotonado intermediário e do nitrogênio imidazol da histidina. Motivos ATCUN são encontrados em várias proteínas biologicamente importantes, incluindo albumina sérica humana (a principal proteína de transporte de cobre no plasma). GHK pode ser compreendido como o peptídeo do motivo ATCUN mínimo — o menor fragmento capaz de alcançar esta coordenação de cobre de três nitrogênios.[7]
Base Estrutural para Vias de Degradação: Mecanismos Moleculares
Fotodegradação: Sensibilidade à Luz
A sensibilidade à luz do GHK-Cu surge diretamente de sua coordenação de cobre. A absorção de luz UV e visível pelo centro de cobre(II) pode gerar estados eletrônicos excitados que catalisam reações radicais dentro da cadeia peptídica, particularmente oxidação do anel imidazol da histidina (que é o local mais rico em elétrons no complexo). O centro de cobre atua como fotossensibilizador, convertendo energia de luz absorvida em dano oxidativo — um processo que não ocorre no peptídeo livre de cobre. Isto explica por que GHK-Cu requer proteção contra luz durante o armazenamento.[8]
Degradação Térmica: Efeitos da Temperatura
Temperaturas elevadas aceleram a oxidação catalisada por cobre da cadeia peptídica. O centro de cobre, através de sua capacidade de ciclagem redox, pode gerar espécies reativas de oxigênio a partir do oxigênio molecular dissolvido em taxas que aumentam com a temperatura. O resíduo de histidina é novamente o alvo primário, com produtos de oxidação incluindo 2-oxo-histidina (um marcador bem caracterizado da oxidação de peptídeos mediada por cobre). Armazenamento em temperaturas refrigeradas (2–8°C) ou abaixo minimiza esta via de degradação.[8]
Degradação Induzida por pH: Correlatos Visuais
Como descrito acima, pH ácido causa dissociação do cobre do peptídeo, liberando cobre(II) livre que pode iniciar química radical descontrolada. O correlato visual é uma mudança de cor do azul para o verde — a cor verde surgindo de íons aqua de cobre(II) livres ou espécies de cobre parcialmente coordenadas. pH alcalino pode promover hidrólise da ligação peptídica, particularmente na ligação Gly-His onde o nitrogênio da amida desprotonado é mais suscetível ao ataque nucleofílico. Manter soluções de GHK-Cu dentro da faixa de pH 5,0–6,5 fornece estabilidade ótima.[3]
Implicações Práticas: Traduzindo Estrutura em Protocolo
Requisitos de Reconstituição
Para pesquisadores, as propriedades estruturais do GHK-Cu se traduzem em vários requisitos práticos. A coordenação do cobre determina que a reconstituição deve usar solventes que mantenham pH dentro da faixa 5,0–7,4 — água bacteriostática (pH aproximadamente 5,5) é bem adequada, enquanto diluentes ácidos ou solução salina normal (cujos íons cloreto podem interferir com a coordenação do cobre) devem ser evitados. A fotossensibilidade necessita proteção contra luz através de frascos âmbar, envolvimento com papel alumínio ou armazenamento em ambientes escuros.[8]
A sensibilidade à temperatura requer armazenamento refrigerado de soluções reconstituídas. E o indicador visual de qualidade — a cor azul real — deve ser monitorado rotineiramente como uma avaliação de primeira linha da integridade do complexo. Estas considerações estruturais também informam o desenho experimental: pesquisadores devem verificar que seus tampões experimentais, meios e condições mantêm a integridade estrutural do GHK-Cu durante todo o período de exposição, pois material degradado pode exibir atividade biológica alterada ou diminuída.
Controle de Qualidade Baseado na Estrutura
O monitoramento da integridade estrutural do GHK-Cu em laboratórios de pesquisa deve incorporar tanto avaliações visuais quanto verificações de pH. A cor azul real característica serve como um indicador imediato de coordenação adequada do cobre — qualquer desvio para verde, marrom ou incolor indica compromisso estrutural. Medições de pH devem confirmar que as soluções permanecem dentro da janela funcional de 5,0–7,4, e o armazenamento deve proteger contra luz, calor e extremos de pH.
Para orientação prática abrangente, consulte nosso guia de manuseio e armazenamento de GHK-Cu, e para princípios gerais de avaliação de qualidade de peptídeos, veja nosso artigo sobre pureza peptídica em estudos científicos. Estas considerações estruturais fundamentam não apenas a compreensão teórica do GHK-Cu, mas também sua aplicação prática eficaz em ambientes de pesquisa, garantindo que a arquitetura molecular única que confere suas propriedades biológicas extraordinárias seja preservada desde a síntese até a aplicação experimental.