Cadre théorique de la chimie de coordination peptide-métal
Il a été démontré que l'activité biologique du complexe GHK-Cu repose entièrement sur ses propriétés de coordination moléculaire — particulièrement l'organisation de trois acides aminés autour d'un ion cuivre(II) central pour créer un complexe aux propriétés biologiques remarquablement diversifiées. La chimie de coordination du GHK-Cu détermine sa stabilité thermodynamique, sa capacité de transport du cuivre, ses interactions avec les récepteurs cellulaires, et ultimement, sa capacité à moduler plus de 4 000 gènes humains.[1]
Cette analyse structurale détaillée s'adresse aux chercheurs désireux de comprendre les bases chimiques des propriétés biologiques du GHK-Cu à des fins de recherche en laboratoire. Pour contextualiser les mécanismes biologiques rendus possibles par cette structure, consultez notre article sur les mécanismes d'action. Pour les implications pratiques des propriétés structurales du GHK-Cu — notamment sa photosensibilité, sa dépendance au pH et ses voies de dégradation — référez-vous à notre guide de manipulation et stockage.
Principes fondamentaux de coordination ATCUN
Le GHK appartient à une classe structurale plus large connue sous le nom de motifs ATCUN (amino-terminal copper and nickel binding) — séquences présentant la formule générale X-X-His (où X représente tout acide aminé), qui coordonnent le cuivre(II) via l'azote amino N-terminal, l'azote déprotoné de la liaison peptidique intermédiaire, et l'azote imidazole de l'histidine. Ces motifs ATCUN se retrouvent dans plusieurs protéines biologiquement importantes, notamment l'albumine sérique humaine (principale protéine de transport du cuivre dans le plasma) et l'extrémité N-terminale de l'histone H2B. Il a été établi que le GHK constitue le peptide motif ATCUN minimal — le plus petit fragment capable d'atteindre cette coordination tri-azotée du cuivre et les activités biologiques qu'elle permet.[7]
Architecture moléculaire du squelette tripeptidique
Caractérisation séquentielle et propriétés chimiques
Le GHK se compose de trois acides aminés liés par des liaisons peptidiques standard : glycine (Gly, position 1), L-histidine (His, position 2), et L-lysine (Lys, position 3). La formule moléculaire du peptide libre de cuivre est C12H22N6O4, tandis que le complexe cuivrique complet présente la formule C14H23CuN6O4 et un poids moléculaire de 401,91 g/mol. Le composé est enregistré sous le numéro CAS 89030-95-5 et PubChem CID 73587.[2]
Chaque résidu contribue des propriétés distinctes au complexe. La glycine, le plus petit acide aminé dépourvu de chaîne latérale, confère une flexibilité conformationnelle à l'extrémité N-terminale et contribue à la coordination du cuivre via son groupe alpha-aminé et l'azote de sa liaison peptidique. Le cycle imidazole de l'histidine constitue le site de liaison primaire du cuivre et la pierre angulaire structurale de l'ensemble du complexe — les atomes d'azote imidazole figurent parmi les donneurs biologiques les plus puissants pour le cuivre(II), et les résidus histidine servent de ligands cuivriques dans de nombreuses métalloenzymes incluant la superoxyde dismutase et la cytochrome c oxydase. La lysine fournit un groupe epsilon-aminé positivement chargé à l'extrémité de sa chaîne latérale, qui contribue à la solubilité aqueuse du peptide, participe aux interactions électrostatiques avec les surfaces biologiques anioniques, et peut contribuer à la coordination intermoléculaire dans les formes cristallines du complexe.[3]
Rationalisation de la séquence spécifique
La séquence Gly-His-Lys n'est pas arbitraire — elle est optimisée pour la liaison cuivrique. L'histidine en position 2 est critique car elle place le cycle imidazole adjacent au groupe amino N-terminal, créant une poche de chélation idéalement dimensionnée pour le cuivre(II). Il a été observé que les peptides avec l'histidine en position 3 ou plus éloignée du terminus présentent une affinité cuivrique dramatiquement réduite. La glycine en position 1 est importante car l'absence de chaîne latérale minimise l'interférence stérique avec la sphère de coordination du cuivre. Le remplacement de la glycine par des acides aminés plus volumineux réduit l'affinité de liaison et altère la géométrie du complexe résultant.[3]
Géométrie de coordination du cuivre(II)
Sphère de coordination primaire
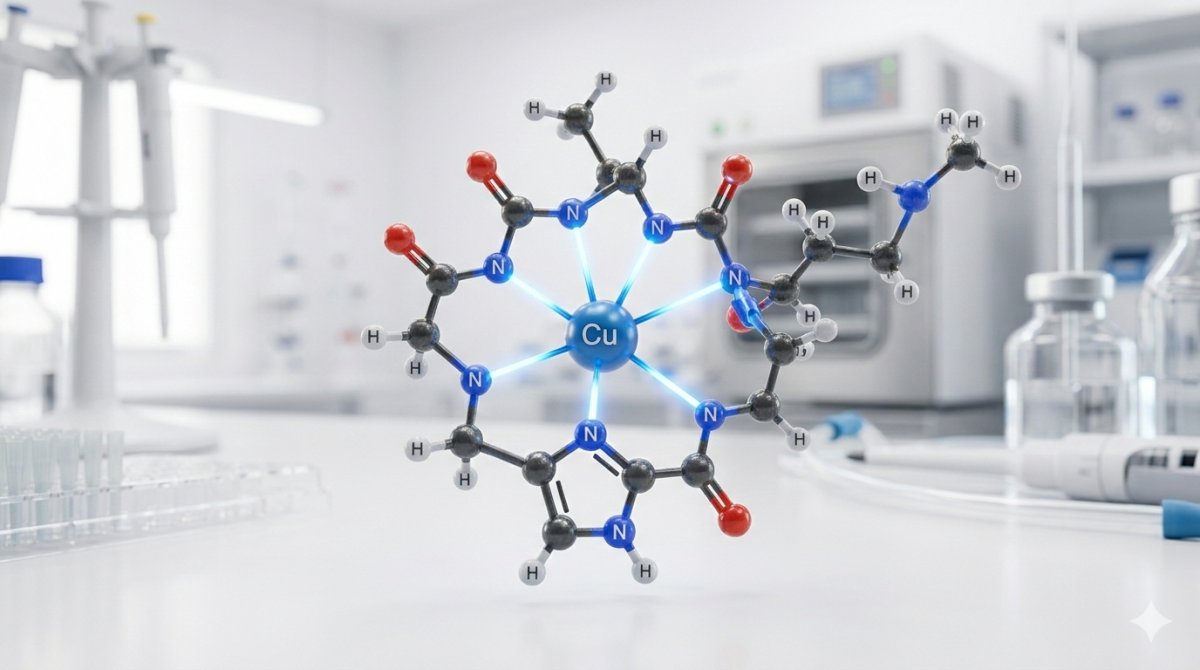
Dans le complexe GHK-Cu, l'ion cuivre(II) est coordonné par trois atomes donneurs d'azote du squelette peptidique, formant la sphère de coordination primaire. Le premier donneur est l'azote Nδ1 ou Nε2 du cycle imidazole de l'histidine (le donneur individuel le plus puissant du complexe). Le second est l'azote alpha-aminé du résidu glycine à l'extrémité N-terminale. Le troisième est l'azote déprotoné de la liaison peptidique (azote amide) entre la glycine et l'histidine — une coordination qui nécessite le déplacement du proton amide et caractérise les complexes cuivre(II)-peptide de forte affinité.[4]
Cette coordination tri-azotée crée un chélate hautement stable avec l'ion cuivre lié au sein de deux cycles fusionnés à cinq membres : un cycle comprenant Cu–NH₂(Gly)–CH₂–CO–N⁻ (le groupe amino glycine à travers la liaison peptidique déprotonée) et un second cycle impliquant l'azote imidazole de l'histidine. La géométrie à cinq membres est thermodynamiquement favorable pour la coordination du cuivre(II), contribuant à la constante de stabilité remarquablement élevée du complexe.[4]
Coordination étendue et structure à l'état solide
L'analyse cristallographique par diffraction des rayons X a révélé qu'à l'état solide, la sphère de coordination du cuivre s'étend au-delà des trois donneurs azotés peptidiques. L'oxygène carboxyle du résidu lysine d'une molécule GHK-Cu voisine occupe une position de coordination axiale, créant une géométrie pyramidale carrée étendue (quatre donneurs équatoriaux plus un donneur axial). Cette coordination intermoléculaire produit une structure de chaîne polymérique dans les cristaux, où les unités GHK-Cu adjacentes sont liées par des ponts cuivre-carboxylate.[4]
En solution, la quatrième position équatoriale est occupée par une molécule d'eau ou un autre donneur disponible (tel que le carboxylate d'un complexe voisin à des concentrations plus élevées), et la géométrie fluctue entre plan carré et pyramidal carré selon la disponibilité des ligands axiaux. Cette flexibilité structurale en solution peut être fonctionnellement significative — permettant au GHK-Cu d'adapter sa sphère de coordination à différents environnements cellulaires et partenaires de liaison.[5]
Constante de stabilité et affinité cuivrique
La stabilité du complexe GHK-Cu est quantifiée par sa constante de stabilité log₁₀ (log K) de 16,44 — une valeur exceptionnellement élevée pour un complexe tripeptide-cuivre. À titre comparatif, l'acide aminé histidine seul lie le cuivre(II) avec un log K d'environ 10,2, et la plupart des complexes dipeptide-cuivre présentent des valeurs log K entre 8 et 14. La constante de stabilité du GHK de 16,44 reflète la contribution coopérative de trois donneurs azotés dans un arrangement géométrique optimal, créant une poche de chélation thermodynamiquement préorganisée pour la liaison cuivre(II).[3]
Cette haute affinité présente des implications biologiques importantes. Aux concentrations physiologiques de cuivre, l'équilibre favorise fortement la formation du complexe — essentiellement tout GHK en présence de cuivre(II) suffisant existera sous forme GHK-Cu. Cependant, l'affinité n'est pas si extrême qu'elle empêche la libération de cuivre vers des accepteurs biologiques de plus haute affinité tels que l'albumine, la céruloplasmine, ou les chaperons cuivriques intracellulaires. Cette affinité intermédiaire — suffisamment forte pour lier et transporter le cuivre, mais pas assez forte pour le séquestrer irréversiblement — positionne le GHK-Cu comme un agent de livraison de cuivre efficace plutôt que simplement un chélateur cuivrique.[1]
Formation de complexes binaires et ternaires
Le complexe binaire : GHK-Cu
L'espèce GHK-cuivre la plus simple est le complexe binaire 1:1 (GHK-Cu), dans lequel une molécule peptidique coordonne un ion cuivre(II) via les trois donneurs azotés décrits précédemment. Cette espèce prédomine au pH physiologique (7,4) lorsque GHK et cuivre sont présents en ratios équimolaires ou quasi-équimolaires. Le complexe binaire est la forme la plus extensivement caractérisée spectroscopiquement et constitue l'espèce le plus communément sous-entendue par la désignation "GHK-Cu" dans la littérature de recherche.[5]
Le complexe ternaire : (GHK)₂-Cu
Lorsque GHK est présent en excès relatif par rapport au cuivre, un complexe ternaire 2:1 peut se former — désigné (GHK)₂-Cu — dans lequel deux molécules peptidiques GHK coordonnent le même ion cuivre(II). Dans cette espèce, chaque molécule peptidique contribue son azote imidazole histidine et un ou deux donneurs additionnels, avec le cuivre coordonné dans une géométrie distordue qui accommode les ligands des deux chaînes peptidiques. Le complexe ternaire a été détecté par méthodes spectroscopiques et peut représenter une fraction significative des espèces GHK-cuivre totales dans des conditions où la concentration peptidique excède substantiellement la concentration cuivrique.[5]
La pertinence biologique du complexe ternaire constitue un domaine d'investigation actif. Des études sur la pénétration cutanée ont démontré que (GHK)₂-Cu peut traverser le stratum corneum, suggérant que les formes binaire et ternaire peuvent contribuer aux effets topiques du GHK-Cu. La question de savoir si les deux formes présentent une activité biologique équivalente ou affichent des profils fonctionnels distincts demeure incomplètement résolue.[6]
Comportement dépendant du pH
Coordination cuivrique en fonction du pH
La chimie de coordination du GHK-Cu est fortement dépendante du pH, propriété ayant des implications pratiques directes pour la manipulation, la formulation, et la conception expérimentale. Aux valeurs de pH acides (en dessous d'environ 4,5), la protonation des donneurs azotés amino et imidazole perturbe la coordination cuivrique, conduisant à la dissociation du complexe et à la libération d'ions cuivre(II) libres. Le cuivre libre n'est plus chélaté sous forme biologiquement contrôlée et peut catalyser la génération de radicaux de type Fenton, contribuant à la dégradation oxydative du peptide et compliquant potentiellement les résultats expérimentaux.[3]
Au pH légèrement acide à neutre (5,0–7,4), les trois donneurs azotés sont progressivement déprotonés et la coordination cuivrique est stable. Cette gamme représente la fenêtre de pH fonctionnelle pour le GHK-Cu et correspond à la gamme de stabilité optimale pour la manipulation et le stockage. Le complexe atteint sa stabilité maximale autour du pH 7,4 (pH physiologique), où les trois donneurs azotés sont pleinement engagés avec le centre cuivrique.[3]
Au pH alcalin (au-dessus de 8,5), des réactions concurrentielles peuvent survenir, incluant la formation d'hydroxyde de cuivre et l'hydrolyse potentielle du squelette peptidique. Bien que le complexe demeure largement intact au pH légèrement alcalin, les conditions fortement basiques devraient être évitées dans les formulations de recherche. Ces considérations de pH sont discutées dans un contexte pratique dans notre guide de manipulation et stockage.
Méthodologies de caractérisation spectroscopique
Spectroscopie UV-visible
La caractéristique la plus immédiatement reconnaissable du GHK-Cu correctement formé est sa couleur bleue royale distinctive, résultant des transitions électroniques d-d de l'ion cuivre(II) au sein de sa sphère de coordination à donneurs azotés. Le spectre d'absorption UV-visible affiche une bande d'absorption large centrée près de 600 nm (la bande de transition d-d caractéristique du cuivre(II) dans un champ de ligands riche en azote) et une bande de transfert de charge plus intense dans la région UV proche. Cette couleur bleue sert d'indicateur pratique de contrôle qualité : le GHK-Cu correctement reconstitué devrait produire une solution bleue claire, et les déviations — vert (indiquant une dissociation cuivrique partielle), brun (indiquant une oxydation), ou incolore (indiquant une perte cuivrique complète) — signalent une dégradation.[4]
Résonance paramagnétique électronique (RPE)
La spectroscopie RPE fournit des informations détaillées sur l'environnement de coordination du cuivre dans le GHK-Cu. Le cuivre(II) est un ion paramagnétique d⁹, et son spectre RPE est sensible au nombre, type, et géométrie des ligands coordinants. Le GHK-Cu affiche des paramètres RPE (valeurs g et constantes de couplage hyperfin) cohérents avec une géométrie de coordination plan carré ou tétragonalement distordue avec des donneurs principalement azotés — confirmant le modèle de coordination tri-azotée dérivé des données cristallographiques. La RPE s'est révélée particulièrement précieuse pour étudier les changements de coordination dépendants du pH, car les paramètres spectraux se décalent de manière mesurable lorsque les donneurs sont protonés ou substitués.[5]
Résonance magnétique nucléaire (RMN)
Bien que le cuivre(II) soit paramagnétique et élargisse les signaux RMN proches (rendant difficile l'observation directe des noyaux liés au cuivre), la spectroscopie RMN d'analogues zinc(II) diamagnétiques et la comparaison des spectres GHK libre de cuivre avec GHK-Cu ont fourni des informations sur les changements conformationnels induits par la liaison métallique. Ces études confirment que la coordination cuivrique contraint le squelette peptidique, réduisant la flexibilité conformationnelle du segment Gly-His et positionnant la chaîne latérale lysine dans une orientation spécifique relative au centre métallique.[5]
Spectroscopie d'absorption des rayons X
La spectroscopie de structure près du seuil d'absorption des rayons X (XANES) et de structure fine d'absorption étendue des rayons X (EXAFS) au seuil K du cuivre fournissent des informations structurelles spécifiques à l'élément sur le centre cuivrique dans le GHK-Cu — incluant le nombre de coordination, les types d'atomes donneurs, et les distances de liaison — dans les échantillons en solution et à l'état solide. L'analyse EXAFS confirme des distances de liaison cuivre-azote d'environ 1,95–2,00 Å dans le plan équatorial, cohérentes avec le modèle cristallographique de coordination tri-azotée.[4]
Relations structure-activité selon la pathologie
Rôle essentiel du cuivre dans l'activité biologique
Les études comparatives entre le GHK-Cu complexé au cuivre et le peptide GHK libre de cuivre démontrent de manière consistante que le complexe complet présente une activité biologique substantiellement supérieure sur virtuellement tous les paramètres mesurés. Cette dépendance au cuivre est mécanistiquement logique : l'activité antioxydante de type SOD requiert la capacité de cyclage redox du centre cuivrique (Cu²⁺ ↔ Cu⁺) ; la livraison de cuivre aux métalloenzymes nécessite du cuivre biodisponible ; et les contraintes conformationnelles imposées par le cuivre sur le squelette peptidique peuvent être nécessaires pour des interactions productives avec les récepteurs.[1]
Modifications séquentielles et études d'analogues
Les études de relations structure-activité utilisant des analogues GHK ont illuminé quelles caractéristiques structurales sont essentielles pour la fonction biologique. Le remplacement de l'histidine par des acides aminés non-coordinants abolit la plupart de l'activité biologique, confirmant la centralité de la coordination imidazole-cuivre. La substitution de la glycine en position 1 par des acides aminés plus larges réduit l'affinité cuivrique et atténue l'activité, démontrant l'importance d'une interférence stérique minimale au site de coordination N-terminal. La modification de la chaîne latérale lysine — par acétylation du groupe ε-aminé ou remplacement par l'arginine — produit des effets plus nuancés qui varient selon le paramètre, suggérant que la lysine contribue aux interactions avec les récepteurs et à la capture cellulaire par des mécanismes partiellement indépendants de la coordination cuivrique.[3]
Bases structurales des voies de dégradation
Photodégradation
La sensibilité à la lumière du GHK-Cu découle directement de sa coordination cuivrique. L'absorption de lumière UV et visible par le centre cuivre(II) peut générer des états électroniques excités qui catalysent les réactions radicalaires au sein du squelette peptidique, particulièrement l'oxydation du cycle imidazole de l'histidine (qui est le site le plus riche en électrons du complexe). Le centre cuivrique agit comme photosensibilisateur, convertissant l'énergie lumineuse absorbée en dommages oxydatifs — un processus qui ne survient pas dans le peptide libre de cuivre. Ceci explique pourquoi le GHK-Cu nécessite une protection lumineuse durant le stockage, tandis que de nombreux peptides libres de cuivre n'en nécessitent pas.[8]
Dégradation thermique
Les températures élevées accélèrent l'oxydation catalysée par le cuivre du squelette peptidique. Le centre cuivrique, par sa capacité de cyclage redox, peut générer des espèces réactives de l'oxygène à partir de l'oxygène moléculaire dissous à des taux qui augmentent avec la température. Le résidu histidine est à nouveau la cible primaire, avec des produits d'oxydation incluant la 2-oxo-histidine (marqueur bien caractérisé de l'oxydation peptidique médiée par le cuivre). Le stockage à températures réfrigérées (2–8°C) ou inférieures minimise cette voie de dégradation.[8]
Dégradation induite par le pH
Comme décrit précédemment, le pH acide cause la dissociation du cuivre du peptide, libérant du cuivre(II) libre qui peut initier une chimie radicalaire incontrôlée. Le corrélat visuel est un changement de couleur du bleu vers le vert — la couleur verte résultant des ions aqua cuivre(II) libres ou d'espèces cuivriques partiellement coordonnées. Le pH alcalin peut promouvoir l'hydrolyse des liaisons peptidiques, particulièrement à la liaison Gly-His où l'azote amide déprotoné est le plus susceptible à l'attaque nucléophile. Maintenir les solutions GHK-Cu dans la gamme de pH 5,0–6,5 fournit une stabilité optimale qui équilibre l'intégrité de la coordination cuivrique contre la susceptibilité hydrolytique.[3]
Implications méthodologiques des propriétés structurales
Pour les chercheurs destinant ce composé à un usage en laboratoire, les propriétés structurales du GHK-Cu se traduisent par plusieurs exigences pratiques. La coordination cuivrique dicte que la reconstitution doit utiliser des solvants maintenant le pH dans la gamme 5,0–7,4 — l'eau bactériostatique (pH environ 5,5) est bien adaptée, tandis que les diluants acides ou le sérum physiologique normal (dont les ions chlorure peuvent interférer avec la coordination cuivrique) devraient être évités. La photosensibilité nécessite une protection lumineuse par flacons ambrés, emballage papier aluminium, ou stockage en environnements sombres. La sensibilité thermique requiert un stockage réfrigéré des solutions reconstituées. Et l'indicateur qualitatif visuel — la couleur bleue royale — devrait être routinièrement surveillé comme évaluation de première ligne de l'intégrité du complexe.[8]
Ces considérations structurales informent également la conception expérimentale : les chercheurs devraient vérifier que leurs tampons expérimentaux, milieux, et conditions maintiennent l'intégrité structurale du GHK-Cu tout au long de la période d'exposition, car le matériel dégradé peut présenter une activité biologique altérée ou diminuée. Pour des conseils pratiques compréhensifs destinés à un usage en laboratoire de recherche, consultez notre guide de manipulation et stockage GHK-Cu, et pour les principes généraux d'évaluation qualitative peptidique, voir notre article sur la pureté peptidique dans les études scientifiques.