Relevancia Clínica de la Estructura Molecular
La comprensión de la estructura molecular del complejo GHK-Cu resulta fundamental para interpretar sus propiedades biológicas observadas en estudios de investigación. El tripéptido glicina-histidina-lisina coordinado con un ion cobre(II) presenta una arquitectura molecular específica que determina su capacidad para modular más de 4,000 genes humanos, su actividad antioxidante tipo superóxido dismutasa, y sus interacciones con receptores celulares.[1]
Esta configuración estructural única explica por qué el complejo GHK-Cu exhibe actividad biológica significativamente superior comparado con el péptido libre de cobre. La geometría de coordinación específica del ion metálico con los tres residuos aminoacídicos crea un sistema molecular optimizado para el transporte de cobre bioactivo y la modulación de procesos celulares regenerativos. Para información sobre los mecanismos biológicos habilitados por esta estructura, véase nuestro artículo sobre mecanismo de acción del GHK-Cu.
Caracterización del Esqueleto Peptídico: Secuencia Gly-His-Lys
Propiedades Moleculares de la Secuencia Aminoacídica
El tripéptido GHK está constituido por tres residuos aminoacídicos unidos mediante enlaces peptídicos convencionales: glicina (Gly, posición 1), L-histidina (His, posición 2) y L-lisina (Lys, posición 3). La fórmula molecular del péptido libre de cobre es C12H22N6O4, mientras que el complejo completo con cobre presenta la fórmula C14H23CuN6O4 y un peso molecular de 401.91 g/mol. El compuesto se encuentra registrado bajo el número CAS 89030-95-5 y PubChem CID 73587.[2]
Cada residuo aminoacídico contribuye propiedades específicas al complejo molecular. La glicina, el aminoácido más pequeño sin cadena lateral, proporciona flexibilidad conformacional en el extremo N-terminal y contribuye tanto su grupo alfa-amino como el nitrógeno del enlace peptídico a la coordinación del cobre. El anillo imidazol de la histidina constituye el sitio primario de unión al cobre y representa la piedra angular estructural del complejo completo: los átomos de nitrógeno del imidazol se encuentran entre los donadores biológicos más fuertes para el cobre(II), y los residuos de histidina sirven como ligandos de cobre en numerosas metaloenzimas incluyendo superóxido dismutasa y citocromo c oxidasa.[3]
La lisina aporta un grupo épsilon-amino cargado positivamente en el extremo de su cadena lateral, el cual contribuye a la solubilidad acuosa del péptido, participa en interacciones electrostáticas con superficies biológicas aniónicas, y puede contribuir a la coordinación intermolecular en formas cristalinas del complejo. Esta diversidad funcional de los tres residuos crea un sistema molecular optimizado para la coordinación metálica y la actividad biológica.[3]
Optimización Estructural de la Secuencia
La secuencia Gly-His-Lys no es arbitraria sino que está optimizada para la unión al cobre. La histidina en posición 2 resulta crítica porque posiciona el anillo imidazol adyacente al grupo amino N-terminal, creando una cavidad de quelación idealmente dimensionada para el cobre(II). Péptidos con histidina en posición 3 o más alejada del extremo terminal exhiben afinidad por el cobre dramáticamente reducida. La glicina en posición 1 es importante porque su carencia de cadena lateral minimiza la interferencia estérica con la esfera de coordinación del cobre. La sustitución de glicina con aminoácidos más voluminosos reduce la afinidad de unión y altera la geometría del complejo resultante.[3]
Geometría de Coordinación del Cobre(II)
Esfera de Coordinación Primaria
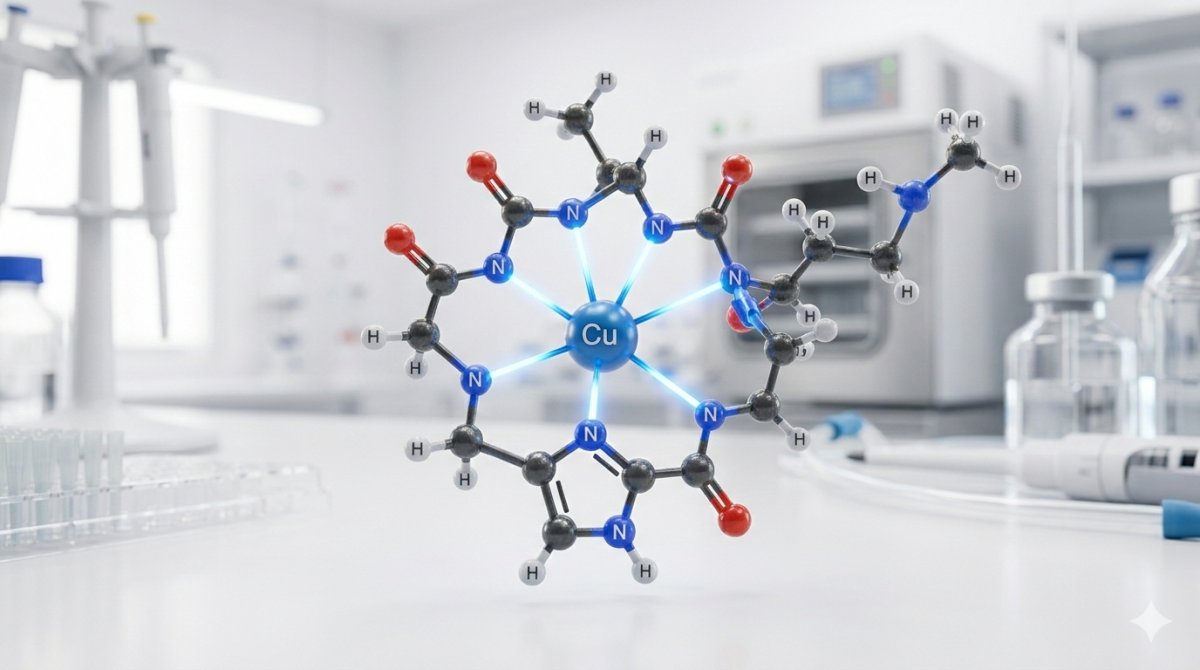
En el complejo GHK-Cu, el ion cobre(II) se coordina mediante tres átomos de nitrógeno donadores del esqueleto peptídico, formando la esfera de coordinación primaria. El primer donador es el nitrógeno Nδ1 o Nε2 del anillo imidazol de la histidina (el donador individual más fuerte en el complejo). El segundo es el nitrógeno alfa-amino del residuo de glicina en el extremo N-terminal. El tercero es el nitrógeno del enlace peptídico desprotonado (nitrógeno amida) entre glicina e histidina, una coordinación que requiere el desplazamiento del protón amida y es característica de complejos cobre(II)-péptido de alta estabilidad.[4]
Esta coordinación triple de nitrógenos crea un quelato altamente estable con el ion cobre unido dentro de dos anillos de cinco miembros fusionados: un anillo que comprende Cu–NH₂(Gly)–CH₂–CO–N⁻ (el grupo amino de glicina a través del enlace peptídico desprotonado) y un segundo anillo que involucra el nitrógeno imidazol de la histidina. La geometría de anillo de cinco miembros es termodinámicamente favorable para la coordinación del cobre(II), contribuyendo a la constante de estabilidad notablemente alta del complejo.[4]
Coordinación Extendida y Estructura en Estado Sólido
El análisis cristalográfico de rayos X ha revelado que en estado sólido, la esfera de coordinación del cobre se extiende más allá de los tres donadores de nitrógeno peptídicos. El oxígeno carboxílico del residuo de lisina de una molécula vecina de GHK-Cu ocupa una posición de coordinación axial, creando una geometría piramidal cuadrada extendida (cuatro donadores ecuatoriales más un donador axial). Esta coordinación intermolecular produce una estructura de cadena polimérica en cristales, donde unidades adyacentes de GHK-Cu se enlazan a través de puentes cobre-carboxilato.[4]
En solución, la cuarta posición ecuatorial es ocupada por una molécula de agua u otro donador disponible (como el carboxilato de un complejo vecino a concentraciones más altas), y la geometría fluctúa entre planar cuadrada y piramidal cuadrada dependiendo de la disponibilidad de ligandos axiales. Esta flexibilidad estructural en solución puede ser funcionalmente significativa, permitiendo que el GHK-Cu adapte su esfera de coordinación a diferentes entornos celulares y compañeros de unión.[5]
Constante de Estabilidad y Afinidad por el Cobre
La estabilidad del complejo GHK-Cu se cuantifica mediante su constante de estabilidad log₁₀ (log K) de 16.44, un valor excepcionalmente alto para un complejo tripéptido-cobre. Para comparación, el aminoácido histidina solo une cobre(II) con un log K de aproximadamente 10.2, y la mayoría de complejos dipéptido-cobre exhiben valores de log K entre 8 y 14. La constante de estabilidad del GHK de 16.44 refleja la contribución cooperativa de tres donadores de nitrógeno en un arreglo geométrico óptimo, creando una cavidad de quelación que está termodinámicamente preorganizada para la unión del cobre(II).[3]
Esta alta afinidad tiene implicaciones biológicas importantes. A concentraciones fisiológicas de cobre, el equilibrio favorece fuertemente la formación del complejo: esencialmente todo el GHK en presencia de suficiente cobre(II) existirá como GHK-Cu. Sin embargo, la afinidad no es tan extrema como para prevenir la liberación de cobre a aceptores biológicos de mayor afinidad como albúmina, ceruloplasmina, o chaperonas de cobre intracelulares. Esta afinidad intermedia, suficientemente fuerte para unir y transportar cobre, pero no tan fuerte como para secuestrarlo irreversiblemente, posiciona al GHK-Cu como un agente efectivo de entrega de cobre más que simplemente un quelante de cobre.[1]
Formación de Complejos Binarios y Ternarios
El Complejo Binario: GHK-Cu
La especie más simple GHK-cobre es el complejo binario 1:1 (GHK-Cu), en el cual una molécula peptídica coordina un ion cobre(II) a través de los tres donadores de nitrógeno descritos anteriormente. Esta especie predomina a pH fisiológico (7.4) cuando GHK y cobre están presentes en relaciones equimolares o cerca de equimolares. El complejo binario es la forma más extensamente caracterizada espectroscópicamente y es la especie más comúnmente implicada por la designación "GHK-Cu" en la literatura de investigación.[5]
El Complejo Ternario: (GHK)₂-Cu
Cuando el GHK está presente en exceso relativo al cobre, puede formarse un complejo ternario 2:1, designado (GHK)₂-Cu, en el cual dos moléculas peptídicas de GHK coordinan el mismo ion cobre(II). En esta especie, cada molécula peptídica contribuye su nitrógeno imidazol de histidina y uno o dos donadores adicionales, con el cobre coordinado en una geometría distorsionada que acomoda ligandos de ambas cadenas peptídicas. El complejo ternario ha sido detectado mediante métodos espectroscópicos y puede representar una fracción significativa de las especies totales GHK-cobre bajo condiciones donde la concentración peptídica excede sustancialmente la concentración de cobre.[5]
La relevancia biológica del complejo ternario es un área activa de investigación. Estudios sobre penetración cutánea han demostrado que el (GHK)₂-Cu puede atravesar el estrato córneo, sugiriendo que tanto las formas binaria como ternaria pueden contribuir a los efectos tópicos del GHK-Cu. Si las dos formas exhiben actividad biológica equivalente o muestran perfiles funcionales distintos permanece incompletamente resuelto.[6]
Comportamiento Dependiente del pH
Coordinación del Cobre en Función del pH
La química de coordinación del GHK-Cu es fuertemente dependiente del pH, una propiedad con implicaciones prácticas directas para el manejo, formulación y diseño experimental. A valores de pH ácidos (por debajo de aproximadamente 4.5), la protonación de los donadores de nitrógeno amino e imidazol interrumpe la coordinación del cobre, llevando a la disociación del complejo y liberación de iones cobre(II) libres. El cobre libre ya no está quelado en una forma biológicamente controlada y puede catalizar la generación de radicales tipo Fenton, contribuyendo a la degradación oxidativa del péptido y potencialmente complicando los resultados experimentales.[3]
A pH ligeramente ácido a neutro (5.0–7.4), los tres donadores de nitrógeno se desprotona progresivamente y la coordinación del cobre es estable. Este rango representa la ventana de pH funcional para el GHK-Cu y corresponde al rango óptimo de estabilidad para manejo y almacenamiento. El complejo alcanza máxima estabilidad alrededor del pH 7.4 (pH fisiológico), donde los tres donadores de nitrógeno están completamente comprometidos con el centro de cobre.[3]
A pH alcalino (por encima de 8.5), pueden ocurrir reacciones competitivas, incluyendo formación de hidróxido de cobre e hidrólisis potencial del esqueleto peptídico. Aunque el complejo permanece en gran parte intacto a pH ligeramente alcalino, las condiciones fuertemente básicas deben evitarse en formulaciones de investigación. Para implicaciones prácticas detalladas de estas consideraciones de pH, véase nuestra guía de manejo y almacenamiento.
Caracterización Espectroscópica
Espectroscopía UV-Visible
La característica más inmediatamente reconocible del GHK-Cu correctamente formado es su color azul real distintivo, que surge de las transiciones electrónicas d-d del ion cobre(II) dentro de su esfera de coordinación de donadores de nitrógeno. El espectro de absorción UV-visible muestra una banda de absorción amplia centrada cerca de 600 nm (la banda de transición d-d característica del cobre(II) en un campo de ligandos rico en nitrógeno) y una banda de transferencia de carga más intensa en la región UV cercana. Este color azul sirve como un indicador práctico de control de calidad: el GHK-Cu correctamente reconstituido debe producir una solución azul clara, y las desviaciones verde (indicando disociación parcial del cobre), marrón (indicando oxidación), o incolora (indicando pérdida completa de cobre) señalan degradación.[4]
Resonancia Paramagnética Electrónica (EPR)
La espectroscopía EPR proporciona información detallada sobre el entorno de coordinación del cobre en el GHK-Cu. El cobre(II) es un ion paramagnético d⁹, y su espectro EPR es sensible al número, tipo y geometría de los ligandos coordinantes. El GHK-Cu muestra parámetros EPR (valores g y constantes de acoplamiento hiperfino) consistentes con una geometría de coordinación planar cuadrada o tetragonalmente distorsionada con donadores predominantemente de nitrógeno, confirmando el modelo de coordinación triple de nitrógenos derivado de datos cristalográficos. El EPR ha sido particularmente valioso para estudiar cambios dependientes del pH en la coordinación, ya que los parámetros espectrales cambian mediblemente cuando los donadores se protonan o sustituyen.[5]
Resonancia Magnética Nuclear (RMN)
Aunque el cobre(II) es paramagnético y amplía las señales de RMN cercanas (haciendo difícil la observación directa de núcleos unidos al cobre), la espectroscopía de RMN de análogos diamagnéticos de zinc(II) y la comparación de espectros de GHK libre de cobre con GHK-Cu han proporcionado información sobre cambios conformacionales inducidos por la unión del metal. Estos estudios confirman que la coordinación del cobre restringe el esqueleto peptídico, reduciendo la flexibilidad conformacional del segmento Gly-His y posicionando la cadena lateral de lisina en una orientación específica relativa al centro metálico.[5]
Espectroscopía de Absorción de Rayos X
La estructura cerca del borde de absorción de rayos X (XANES) y la espectroscopía de estructura fina extendida de absorción de rayos X (EXAFS) en el borde K del cobre proporcionan información estructural específica del elemento sobre el centro de cobre en el GHK-Cu, incluyendo número de coordinación, tipos de átomos donadores y distancias de enlace, tanto en solución como en muestras en estado sólido. El análisis EXAFS confirma distancias de enlace cobre-nitrógeno de aproximadamente 1.95–2.00 Å en el plano ecuatorial, consistente con el modelo cristalográfico de coordinación triple de nitrógenos.[4]
Evidencia de Relaciones Estructura-Actividad
El Papel Esencial del Cobre
Estudios comparativos entre el GHK-Cu complejado con cobre y el péptido GHK libre de cobre demuestran consistentemente que el complejo completo exhibe actividad biológica sustancialmente mayor a través de virtualmente todos los endpoints medidos. Esta dependencia del cobre es mecánicamente lógica: la actividad antioxidante tipo SOD requiere la capacidad de ciclado redox del centro de cobre (Cu²⁺ ↔ Cu⁺); la entrega de cobre a metaloenzimas requiere cobre biodisponible; y las restricciones conformacionales impuestas por el cobre sobre el esqueleto peptídico pueden ser necesarias para interacciones productivas con receptores.[1]
Modificaciones de Secuencia y Estudios de Análogos
Los estudios de relación estructura-actividad utilizando análogos de GHK han iluminado qué características estructurales son esenciales para la función biológica. La sustitución de histidina con aminoácidos no coordinantes elimina la mayoría de la actividad biológica, confirmando la centralidad de la coordinación imidazol-cobre. La sustitución de glicina en posición 1 con aminoácidos más grandes reduce la afinidad por el cobre y atenúa la actividad, demostrando la importancia de la interferencia estérica mínima en el sitio de coordinación N-terminal. La modificación de la cadena lateral de lisina a través de acetilación del grupo ε-amino o sustitución con arginina produce efectos más matizados que varían por endpoint, sugiriendo que la lisina contribuye a las interacciones con receptores y captación celular a través de mecanismos parcialmente independientes de la coordinación del cobre.[3]
El Motivo ATCUN
El GHK pertenece a una clase estructural más amplia conocida como péptidos de motivo ATCUN (unión amino-terminal de cobre y níquel), secuencias con la fórmula general X-X-His (donde X es cualquier aminoácido), que coordinan cobre(II) a través del nitrógeno amino N-terminal, el nitrógeno del enlace peptídico intermedio desprotonado, y el nitrógeno imidazol de la histidina. Los motivos ATCUN se encuentran en varias proteínas biológicamente importantes, incluyendo la albúmina sérica humana (la principal proteína transportadora de cobre en plasma) y la cola N-terminal de la histona H2B. El GHK puede entenderse como el péptido de motivo ATCUN mínimo: el fragmento más pequeño capaz de lograr esta coordinación triple de nitrógenos del cobre y las actividades biológicas que habilita.[7]
Base Estructural de las Vías de Degradación
Fotodegradación
La sensibilidad a la luz del GHK-Cu surge directamente de su coordinación con cobre. La absorción de luz UV y visible por el centro de cobre(II) puede generar estados electrónicos excitados que catalizan reacciones de radicales dentro del esqueleto peptídico, particularmente oxidación del anillo imidazol de histidina (que es el sitio más rico en electrones en el complejo). El centro de cobre actúa como un fotosensibilizador, convirtiendo la energía lumínica absorbida en daño oxidativo, un proceso que no ocurre en el péptido libre de cobre. Esto explica por qué el GHK-Cu requiere protección lumínica durante el almacenamiento, mientras que muchos péptidos libres de cobre no la requieren.[8]
Degradación Térmica
Las temperaturas elevadas aceleran la oxidación catalizada por cobre del esqueleto peptídico. El centro de cobre, a través de su capacidad de ciclado redox, puede generar especies reactivas de oxígeno a partir del oxígeno molecular disuelto a velocidades que aumentan con la temperatura. El residuo de histidina es nuevamente el objetivo primario, con productos de oxidación incluyendo 2-oxo-histidina (un marcador bien caracterizado de oxidación peptídica mediada por cobre). El almacenamiento a temperaturas refrigeradas (2–8°C) o por debajo minimiza esta vía de degradación.[8]
Degradación Inducida por pH
Como se describió anteriormente, el pH ácido causa disociación del cobre del péptido, liberando cobre(II) libre que puede iniciar química radical descontrolada. El correlato visual es un cambio de color de azul hacia verde: el color verde surge de iones cobre(II) aqua libres o especies de cobre parcialmente coordinadas. El pH alcalino puede promover hidrólisis del enlace peptídico, particularmente en el enlace Gly-His donde el nitrógeno amida desprotonado es más susceptible al ataque nucleofílico. Mantener las soluciones de GHK-Cu dentro del rango de pH 5.0–6.5 proporciona estabilidad óptima que equilibra la integridad de coordinación del cobre contra la susceptibilidad hidrolítica.[3]
Implicaciones Prácticas de las Propiedades Estructurales para Investigación
Para investigadores, las propiedades estructurales del GHK-Cu se traducen en varios requerimientos prácticos. La coordinación del cobre dicta que la reconstitución debe usar solventes que mantengan el pH dentro del rango 5.0–7.4: el agua bacteriostática (pH aproximadamente 5.5) es bien adecuada, mientras que diluyentes ácidos o solución salina normal (cuyos iones cloruro pueden interferir con la coordinación del cobre) deben evitarse. La fotosensibilidad necesita protección lumínica a través de viales ámbar, envoltura con papel aluminio, o almacenamiento en ambientes oscuros. La sensibilidad a la temperatura requiere almacenamiento refrigerado de soluciones reconstituidas. Y el indicador visual de calidad, el color azul real, debe monitorearse rutinariamente como una evaluación de primera línea de la integridad del complejo.[8]
Estas consideraciones estructurales también informan el diseño experimental: los investigadores deben verificar que sus buffers experimentales, medios y condiciones mantengan la integridad estructural del GHK-Cu a lo largo del período de exposición, ya que el material degradado puede exhibir actividad biológica alterada o disminuida. Para orientación práctica integral, véase nuestra guía de manejo y almacenamiento de GHK-Cu, y para principios generales de evaluación de calidad peptídica, véase nuestro artículo sobre pureza de péptidos en estudios científicos.
Relevancia Estructural para la Investigación Avanzada
El análisis estructural detallado del GHK-Cu proporciona la base molecular necesaria para comprender sus múltiples actividades biológicas observadas en investigación. La coordinación específica del cobre(II) con los tres donadores de nitrógeno del péptido crea un sistema químico único que combina transporte de metal biodisponible, actividad redox controlada, y reconocimiento molecular específico. Esta arquitectura molecular explica la capacidad del complejo para modular procesos celulares fundamentales y justifica su uso continuado como herramienta de investigación en estudios de regeneración celular, únicamente con fines de investigación científica.
La caracterización estructural también informa el desarrollo de análogos y derivados del GHK-Cu para aplicaciones específicas de investigación. La comprensión de qué elementos estructurales son esenciales para la actividad biológica y cuáles pueden modificarse sin pérdida de función permite el diseño racional de nuevos complejos péptido-metal con propiedades optimizadas para estudios experimentales particulares, contribuyendo al avance del conocimiento científico en el campo de la biología molecular y la medicina regenerativa.