Introduction: Why Structure Matters
The biological activity of GHK-Cu cannot be understood without understanding its structure — particularly how three amino acids organize themselves around a single copper(II) ion to create a complex with extraordinarily diverse biological properties. The coordination chemistry of GHK-Cu determines its stability, its copper delivery capacity, its interactions with cellular receptors, and ultimately, its ability to modulate over 4,000 human genes.[1]
This article provides a detailed structural analysis for researchers seeking to understand the chemical basis of GHK-Cu's biological properties. For context on the biological mechanisms enabled by this structure, see our mechanism of action article. For practical implications of GHK-Cu's structural properties — particularly its light sensitivity, pH dependence, and degradation pathways — see our handling and storage guide.
The Tripeptide Backbone: Gly-His-Lys
Amino Acid Sequence and Properties
GHK consists of three amino acids linked by standard peptide bonds: glycine (Gly, position 1), L-histidine (His, position 2), and L-lysine (Lys, position 3). The molecular formula of the copper-free peptide is C12H22N6O4, while the complete copper complex has the formula C14H23CuN6O4 and a molecular weight of 401.91 g/mol. The compound is registered under CAS number 89030-95-5 and PubChem CID 73587.[2]
Each residue contributes distinct properties to the complex. Glycine, the smallest amino acid with no side chain, provides conformational flexibility at the N-terminus and contributes both its alpha-amino group and its peptide bond nitrogen to copper coordination. Histidine's imidazole ring is the primary copper-binding site and the structural cornerstone of the entire complex — imidazole nitrogen atoms are among the strongest biological donors for copper(II), and histidine residues serve as copper ligands in numerous metalloenzymes including superoxide dismutase and cytochrome c oxidase. Lysine provides a positively charged epsilon-amino group at its side chain terminus, which contributes to the peptide's aqueous solubility, participates in electrostatic interactions with anionic biological surfaces, and may contribute to inter-molecular coordination in crystalline forms of the complex.[3]
Why This Particular Sequence
The Gly-His-Lys sequence is not arbitrary — it is optimized for copper binding. The histidine at position 2 is critical because it places the imidazole ring adjacent to the N-terminal amino group, creating a chelation pocket ideally sized for copper(II). Peptides with histidine at position 3 or further from the terminus exhibit dramatically reduced copper affinity. The glycine at position 1 is important because its lack of a side chain minimizes steric interference with the copper coordination sphere. Replacing glycine with bulkier amino acids reduces binding affinity and alters the geometry of the resulting complex.[3]
Copper(II) Coordination Geometry
Primary Coordination Sphere
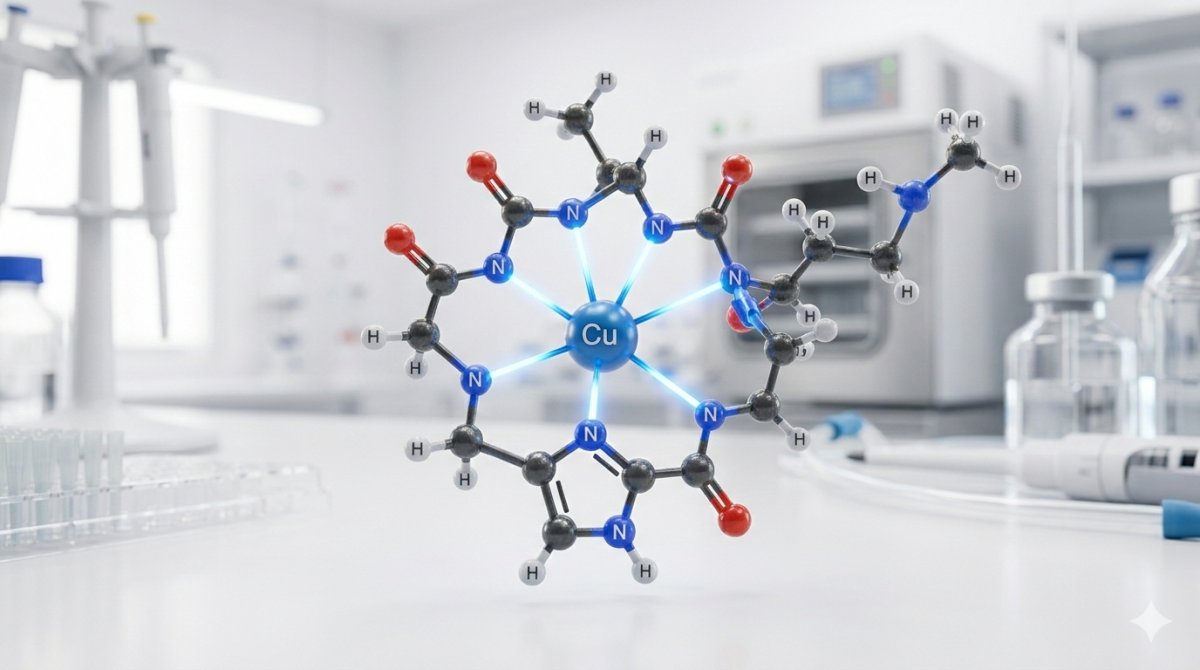
In the GHK-Cu complex, the copper(II) ion is coordinated by three nitrogen donor atoms from the peptide backbone, forming the primary coordination sphere. The first donor is the Nδ1 or Nε2 nitrogen of the histidine imidazole ring (the strongest individual donor in the complex). The second is the alpha-amino nitrogen of the glycine residue at the N-terminus. The third is the deprotonated peptide bond nitrogen (amide nitrogen) between glycine and histidine — a coordination that requires displacement of the amide proton and is characteristic of strong copper(II)-peptide complexes.[4]
This three-nitrogen coordination creates a highly stable chelate with the copper ion bound within two fused five-membered rings: one ring comprising Cu–NH₂(Gly)–CH₂–CO–N⁻ (the glycine amino group through the deprotonated peptide bond) and a second ring involving the histidine imidazole nitrogen. The five-membered ring geometry is thermodynamically favorable for copper(II) coordination, contributing to the complex's remarkably high stability constant.[4]
Extended Coordination and Solid-State Structure
X-ray crystallographic analysis has revealed that in the solid state, the copper coordination sphere extends beyond the three peptide nitrogen donors. The carboxyl oxygen of the lysine residue from a neighboring GHK-Cu molecule occupies an axial coordination position, creating an extended square-pyramidal geometry (four equatorial donors plus one axial donor). This intermolecular coordination produces a polymeric chain structure in crystals, where adjacent GHK-Cu units are linked through copper-carboxylate bridges.[4]
In solution, the fourth equatorial position is occupied by a water molecule or other available donor (such as the carboxylate of a neighboring complex at higher concentrations), and the geometry fluctuates between square-planar and square-pyramidal depending on the availability of axial ligands. This structural flexibility in solution may be functionally significant — allowing GHK-Cu to adapt its coordination sphere to different cellular environments and binding partners.[5]
Stability Constant and Copper Affinity
The stability of the GHK-Cu complex is quantified by its log₁₀ stability constant (log K) of 16.44 — an exceptionally high value for a tripeptide-copper complex. For comparison, the amino acid histidine alone binds copper(II) with a log K of approximately 10.2, and most dipeptide-copper complexes exhibit log K values between 8 and 14. GHK's stability constant of 16.44 reflects the cooperative contribution of three nitrogen donors in an optimal geometric arrangement, creating a chelation pocket that is thermodynamically preorganized for copper(II) binding.[3]
This high affinity has important biological implications. At physiological copper concentrations, the equilibrium strongly favors complex formation — essentially all GHK in the presence of sufficient copper(II) will exist as GHK-Cu. However, the affinity is not so extreme as to prevent copper release to higher-affinity biological acceptors such as albumin, ceruloplasmin, or intracellular copper chaperones. This intermediate affinity — strong enough to bind and transport copper, but not so strong as to sequester it irreversibly — positions GHK-Cu as an effective copper delivery agent rather than simply a copper chelator.[1]
Binary and Ternary Complex Formation
The Binary Complex: GHK-Cu
The simplest GHK-copper species is the 1:1 binary complex (GHK-Cu), in which one peptide molecule coordinates one copper(II) ion through the three nitrogen donors described above. This species predominates at physiological pH (7.4) when GHK and copper are present in equimolar or near-equimolar ratios. The binary complex is the most extensively characterized form spectroscopically and is the species most commonly implied by the designation "GHK-Cu" in the research literature.[5]
The Ternary Complex: (GHK)₂-Cu
When GHK is present in excess relative to copper, a 2:1 ternary complex can form — designated (GHK)₂-Cu — in which two GHK peptide molecules coordinate the same copper(II) ion. In this species, each peptide molecule contributes its histidine imidazole nitrogen and one or two additional donors, with the copper coordinated in a distorted geometry that accommodates ligands from both peptide chains. The ternary complex has been detected by spectroscopic methods and may represent a significant fraction of the total GHK-copper species under conditions where peptide concentration substantially exceeds copper concentration.[5]
The biological relevance of the ternary complex is an active area of investigation. Studies on skin penetration have demonstrated that (GHK)₂-Cu can traverse the stratum corneum, suggesting that both the binary and ternary forms may contribute to topical GHK-Cu effects. Whether the two forms exhibit equivalent biological activity or display distinct functional profiles remains incompletely resolved.[6]
pH-Dependent Behavior
Copper Coordination as a Function of pH
The coordination chemistry of GHK-Cu is strongly pH-dependent, a property with direct practical implications for handling, formulation, and experimental design. At acidic pH values (below approximately 4.5), protonation of the amino and imidazole nitrogen donors disrupts copper coordination, leading to dissociation of the complex and release of free copper(II) ions. The free copper is no longer chelated in a biologically controlled form and can catalyze Fenton-type radical generation, contributing to oxidative degradation of the peptide and potentially complicating experimental results.[3]
At mildly acidic to neutral pH (5.0–7.4), the three nitrogen donors are progressively deprotonated and copper coordination is stable. This range represents the functional pH window for GHK-Cu and corresponds to the optimal stability range for handling and storage. The complex reaches maximum stability around pH 7.4 (physiological pH), where all three nitrogen donors are fully engaged with the copper center.[3]
At alkaline pH (above 8.5), competing reactions can occur, including copper hydroxide formation and potential hydrolysis of the peptide backbone. While the complex remains largely intact at mildly alkaline pH, strongly basic conditions should be avoided in research formulations. These pH considerations are discussed in practical context in our handling and storage guide.
Spectroscopic Characterization
UV-Visible Spectroscopy
The most immediately recognizable feature of properly formed GHK-Cu is its distinctive royal blue color, arising from d-d electronic transitions of the copper(II) ion within its nitrogen-donor coordination sphere. The UV-visible absorption spectrum displays a broad absorption band centered near 600 nm (the d-d transition band characteristic of copper(II) in a nitrogen-rich ligand field) and a more intense charge-transfer band in the near-UV region. This blue color serves as a practical quality control indicator: properly reconstituted GHK-Cu should produce a clear blue solution, and deviations — green (indicating partial copper dissociation), brown (indicating oxidation), or colorlessness (indicating complete copper loss) — signal degradation.[4]
Electron Paramagnetic Resonance (EPR)
EPR spectroscopy provides detailed information about the copper coordination environment in GHK-Cu. Copper(II) is a d⁹ paramagnetic ion, and its EPR spectrum is sensitive to the number, type, and geometry of coordinating ligands. GHK-Cu displays EPR parameters (g-values and hyperfine coupling constants) consistent with a square-planar or tetragonally distorted coordination geometry with predominantly nitrogen donors — confirming the three-nitrogen coordination model derived from crystallographic data. EPR has been particularly valuable for studying pH-dependent changes in coordination, as the spectral parameters shift measurably when donors are protonated or substituted.[5]
Nuclear Magnetic Resonance (NMR)
While copper(II) is paramagnetic and broadens nearby NMR signals (making direct observation of copper-bound nuclei difficult), NMR spectroscopy of diamagnetic zinc(II) analogues and comparison of copper-free GHK with GHK-Cu spectra have provided information about conformational changes induced by metal binding. These studies confirm that copper coordination constrains the peptide backbone, reducing the conformational flexibility of the Gly-His segment and positioning the lysine side chain in a specific orientation relative to the metal center.[5]
X-Ray Absorption Spectroscopy
X-ray absorption near-edge structure (XANES) and extended X-ray absorption fine structure (EXAFS) spectroscopy at the copper K-edge provide element-specific structural information about the copper center in GHK-Cu — including coordination number, donor atom types, and bond distances — in both solution and solid-state samples. EXAFS analysis confirms copper-nitrogen bond distances of approximately 1.95–2.00 Å in the equatorial plane, consistent with the crystallographic model of three-nitrogen coordination.[4]
Structure-Activity Relationships
The Essential Role of Copper
Comparative studies between copper-complexed GHK-Cu and the copper-free peptide GHK consistently demonstrate that the complete complex exhibits substantially greater biological activity across virtually all measured endpoints. This copper dependence is mechanistically logical: the SOD-like antioxidant activity requires the copper center's redox cycling capability (Cu²⁺ ↔ Cu⁺); copper delivery to metalloenzymes requires bioavailable copper; and the copper-imposed conformational constraints on the peptide backbone may be necessary for productive receptor interactions.[1]
Sequence Modifications and Analogue Studies
Structure-activity relationship studies using GHK analogues have illuminated which structural features are essential for biological function. Replacement of histidine with non-coordinating amino acids abolishes most biological activity, confirming the centrality of imidazole-copper coordination. Substitution of glycine at position 1 with larger amino acids reduces copper affinity and attenuates activity, demonstrating the importance of minimal steric interference at the N-terminal coordination site. Modification of the lysine side chain — through acetylation of the ε-amino group or replacement with arginine — produces more nuanced effects that vary by endpoint, suggesting that the lysine contributes to receptor interactions and cellular uptake through mechanisms partially independent of copper coordination.[3]
The ATCUN Motif
GHK belongs to a broader structural class known as ATCUN (amino-terminal copper and nickel binding) motif peptides — sequences with the general formula X-X-His (where X is any amino acid), which coordinate copper(II) through the N-terminal amino nitrogen, the deprotonated intervening peptide bond nitrogen, and the histidine imidazole nitrogen. ATCUN motifs are found in several biologically important proteins, including human serum albumin (the major copper transport protein in plasma) and the histone H2B N-terminal tail. GHK can be understood as the minimal ATCUN motif peptide — the smallest fragment capable of achieving this three-nitrogen copper coordination and the biological activities it enables.[7]
Structural Basis for Degradation Pathways
Photodegradation
GHK-Cu's light sensitivity arises directly from its copper coordination. Absorption of UV and visible light by the copper(II) center can generate excited electronic states that catalyze radical reactions within the peptide backbone, particularly oxidation of the histidine imidazole ring (which is the most electron-rich site in the complex). The copper center acts as a photosensitizer, converting absorbed light energy into oxidative damage — a process that does not occur in the copper-free peptide. This explains why GHK-Cu requires light protection during storage, while many copper-free peptides do not.[8]
Thermal Degradation
Elevated temperatures accelerate copper-catalyzed oxidation of the peptide backbone. The copper center, through its redox cycling capability, can generate reactive oxygen species from dissolved molecular oxygen at rates that increase with temperature. The histidine residue is again the primary target, with oxidation products including 2-oxo-histidine (a well-characterized marker of copper-mediated peptide oxidation). Storage at refrigerated temperatures (2–8°C) or below minimizes this degradation pathway.[8]
pH-Induced Degradation
As described above, acidic pH causes dissociation of the copper from the peptide, releasing free copper(II) that can initiate uncontrolled radical chemistry. The visual correlate is a color shift from blue toward green — the green color arising from free copper(II) aqua ions or partially coordinated copper species. Alkaline pH can promote peptide bond hydrolysis, particularly at the Gly-His bond where the deprotonated amide nitrogen is most susceptible to nucleophilic attack. Maintaining GHK-Cu solutions within the pH 5.0–6.5 range provides optimal stability that balances copper coordination integrity against hydrolytic susceptibility.[3]
Practical Implications of Structural Properties
For researchers, GHK-Cu's structural properties translate into several practical requirements. The copper coordination dictates that reconstitution must use solvents that maintain pH within the 5.0–7.4 range — bacteriostatic water (pH approximately 5.5) is well-suited, while acidic diluents or normal saline (whose chloride ions can interfere with copper coordination) should be avoided. The photosensitivity necessitates light protection through amber vials, aluminum foil wrapping, or storage in dark environments. The temperature sensitivity requires refrigerated storage of reconstituted solutions. And the visual quality indicator — the royal blue color — should be routinely monitored as a first-line assessment of complex integrity.[8]
These structural considerations also inform experimental design: researchers should verify that their experimental buffers, media, and conditions maintain GHK-Cu's structural integrity throughout the exposure period, as degraded material may exhibit altered or diminished biological activity. For comprehensive practical guidance, see our GHK-Cu handling and storage guide, and for general principles of peptide quality assessment, see our article on peptide purity in scientific studies.